Forschung - Generelle Richtung der Forschung in meinem Labor
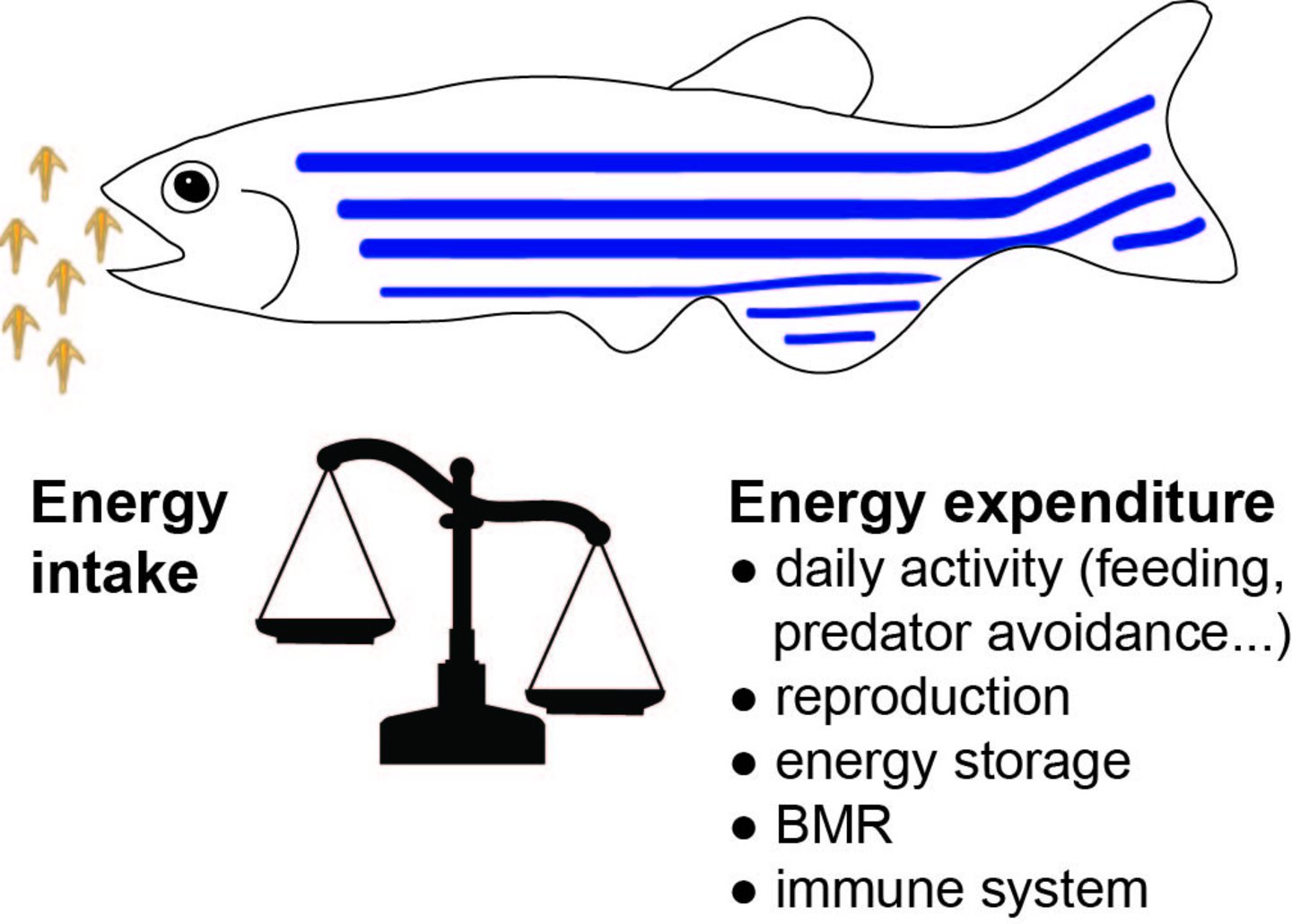
Energiehomöostase ist die Balance zwischen Nahrungsaufnahme und Energieverbrauch. Die Nahrungsaufnahme als input steht in balance zu dem Ruhestoffwechsel (die Energie, die benötigt wird, um den Körper im Ruhezustand zu halten), der thermischen Wirkung von Nahrung (die Energie, die benötigt wird, um Nahrung zu verstoffwechseln und ihre Nährstoffe und Energie verfügbar zu machen) sowie allen aktiven Maßnahmen wie Wachstum, Reproduktion, generelle Physische Aktivität (Nahrungssuche, Sport et cetera).
Wenn dieses System aus dem Gleichgewicht gerät, folgen Stoffwechselkrankheiten, die Mittlerweile eine enorme gesellschaftliche Belastung darstellen. Adipositas, Appetitlosigkeit, Anorexie und Krebs-Kachexie sind alle signifikante Gesellschaftliche Belastungen. Jede dieser Konditionen bringt eine Vielzahl von Komorbiditäten mit sich, vor allem Typ-2-Diabetes mit Fettleibigkeit, aber reproduktive Probleme sind auch komorbid mit beiden Enden der Skala - Fettleibigkeit sowie Anorexie. Daher ist eine gute Balance zwischen Nahrungsaufnahme und -ausgabe entscheidend.
Man könnte postulieren, dass Essen weitgehend eine kulturelle und verhaltensbedingte Belastung - zu viel kalorienreiche Nahrung, zu wenig Bewegung, ob aufgrund von Arbeitszeiten oder verringertem Interesse; und das hat zweifellos Auswirkungen. Unsere Genetik hat jedoch auch einen überraschend großen Einfluss darauf, wie wir unseren Energiehaushalt führen. Eine aktuelle Schätzung von Sir Stephen O'Rahilly lautet, dass "die Genetik die meisten (wahrscheinlich etwa 65%) der Gewichtsschwankungen zwischen Individuen erklärt" (Speakman und O'Rahilly 2012). Die ersten Gene die gefunden wurden stammen aus Mauszuchten der Jackson Laboratories in den sechziger Jahren. Mehrere monogene Mausstämme zeigten Adipositas, aber auch Komorbiditäten wie Diabetes, und diese wurden treffend benannt als 1) obese, 2) diabetic, 3) Agouti (hat auch einen Fellfarbenphänotyp), 4) fatty und 5) tubby Mäuse (Naggert et al., 1997). Die Gene, die diese Phänotypen verursachen, wurden in den neunziger Jahren gefunden und die Forschung auf diesem Gebiet ist seitdem explodiert. Einfach ausgedrückt, das Hormon Leptin wird im Fettgewebe im Verhältnis zur Fettmasse produziert und signalisiert über AgRP- und POMC-Neuronen an Melanocortin-Neuronen, dass genügend Energie in der Peripherie vorhanden ist. Wenn wir fasten, sinkt die Leptinspiegel (von griech. Leptos - dünn) und dies wird durch die Melanocortinneuronen im Hypothalamus des Gehirns als Hunger übersetzt. Mäuse, die zum Beispiel kein Leptin haben (die übergewichtige Mauslinie) sind zutiefst hyperphagisch, weil das Gehirn ständig denkt, dass das Tier verhungert! Folglich sind diese Tiere stark adipös, wobei mehr als die Hälfte des Körpergewichtes aus Fettgewebe besteht. Die Verabreichung von Leptin korrigiert die schwere Fettleibigkeit sowohl bei Mäusen als auch bei Menschen. In einer erstaunlichen Studie fand Sadaf Farooqi ein Mädchen im Alter von 4 Monaten das übermäßig zugenommen hatte und ständig hungrig war. Die Behandlung mit Leptin kurierte die Fettleibigkeit (Farooqi et al. 1999). Während dieser besondere Zustand äußerst selten ist, sind die Kerngene, die an der Homöostase der Maus beteiligt sind, für Menschen konserviert und spielen bei Menschen ähnliche Rollen.
Aber diese Gene sind nicht nur bei Säugetieren, sondern bei allen Wirbeltieren konserviert! Ich arbeite an der Energie-Homöostase im Zebrafisch. Der Zebrafisch ist ein Tier, dessen frühe Entwicklung in den letzten Jahrzehnten intensiv untersucht wurde. Das Genom wurde sequenziert und Labore auf der ganzen Welt haben zu einer beeindruckenden Toolbox beigetragen, um die Zebrafischbiologie zu untersuchen. Wir haben mehrere Mutantenlinien in Leptin, Leptinrezeptor und AgRP sowie verschiedene Markerlinien in der frühen Entwicklung etabliert. Während die Kerngene und -Mechanismen zwischen Wirbeltieren konserviert sind, bestehen wesentliche Unterschiede im Detail. Ein Schwerpunkt des Labors ist es, eine Roadmap zur metabolischen Kontrolle im Zebrafisch zu erstellen, um zu verstehen, welche Aspekte der Energiehomöostase funktionell erhalten sind und welche sich zwischen Spezien unterscheiden.
Der Hauptfokus liegt momentan auf dem Leptin gen bzw dem leptin Rezeptor. Das Hormon wird in Mäusen nur im Fettgewebe exprimiert (Zhang et al. 1994), während es bei Fischen in den meisten bisher untersuchten Arten im Fettgewebe garnicht vorkommt. Wir fanden heraus, dass überfütterte Zebrafische mit Leptin- oder Leptin-Rezeptor-Mutationen im Gegensatz zu Mäusen mit Leptin- oder Leptin-Rezeptor-Mutationen nicht fettleibig werden, ihr Phänotyp liegt jedoch in der Glukoseregulation. Bei Mäusen mit dieser Mutation sind Adipositas und Diabetes komorbid. Das wirft faszinierende Fragen auf - wenn Leptin dem Gehirn nicht signalisiert, was Fett ist, was dann? Was ist der Mechanismus der Glukoregulation durch Leptin bei Fischen und können weitere Erkenntnisse genutzt werden, um zu untersuchen, warum Typ 2 Diabetes bei Menschen manchmal mit Adipositas komorbid ist, aber nicht immer?
Die langfristigen Ziele dieser Forschung sind dreifach: 1) Verbesserung abnormaler Homöostasemuster (mit Auswirkungen auf Tierarten), 2) Einblick in die Evolution der biologischen Vielfalt und 3) Verbesserung des Wachstumsverhaltens von Nutztieren.
Forschung - Spezifische Studien
LepR LOF-Linien – sind sie echt?
Menschen und Mäuse mit einem Verlust in der Leptin Signal Kaskade sind fettleibig. In der Literatur is es unklar, ob Fische mit einem Funktionsverlust von Leptin a oder dem Leptinrezeptor fettleibig sind oder nicht. Es wurde vermutet, dass die Zebrafischlinie Lepr sa1508 möglicherweise hypomorph ist und daher keine Fettleibigkeit aufweist. Wir haben versucht, a) mehr Lepr- und LepA-Allele zu produzieren, in denen dies in unseren Händen getestet werden kann, und b) zu testen, ob die Leptin-Signal Kaskade in unseren Lepr-LOF-Linien funktioniert oder nicht.
Zu diesem Zweck haben wir beschlossen, Leptin zu injizieren und zu prüfen, der Transkribierungsfaktor SOCS3a induziert wird oder nicht. Wir fanden heraus, dass Zebrafischlebern tatsächlich auf IP-Leptin-Injektionen mit einer Hochregulierung von SOCS3a reagieren. Wichtig ist, dass dieser Effekt in allen drei Lepr LOF-Linien, die wir haben, sa1508 und zwei Linien, die wir mit CRISPR Cas9 erstellt haben, gezielt beseitigt wurde. Zumindest was den SOCS-Signalübertragungsweg stromabwärts von Lepr betrifft, handelt es sich bei diesen drei Lepr-LOF-Linien daher um einen echten Funktionsverlust. - (Bagivalu Lakshminarasimha et al., 2021)
Einige Vorbehalte gegenüber Fischwachstumsexperimenten.
Wir haben weiter getestet, ob die Art der Tierhaltung vielleicht Auswirkungen auf das Wachstumsverhalten hat. Häufig werden Zebrafischlinien als -/- x -/- gekreuzt, entweder mit einer Geschwister-Wildtyp-Kreuzung und/oder mit einfachen Wildtyp-Kontrollen, die als unterschiedlicher Bestand gehalten werden. Dies kann als Praxis in Ordnung sein, wenn Mauslinien einer Inzucht verwendet werden, bei denen die genetische Variablität stark reduziert ist. Zebrafische hingegen haben eine hohe Variabilität im Genom, ähnlich wie Menschen. Darüber hinaus hängt das Wachstum von Fischen stark von der Nahrungsmenge und der Dichte der Fische in dem Lebensraum ab, in dem sie sich befinden, sodass selbst geringfügige Unterschiede zwischen den Becken in der Anzahl der Fische oder dem gegebenen Futter zu Wachstumsunterschieden führen können.
Wenn wir für all diese Effekte kontrollieren indem wir eine Het-x-Het-Kreuzung züchteten und die Fische im Alter von 3 Monaten trennten, ihr Gewicht und ihre Länge ermittelten und dann die Fische genotypisierten, stellten wir wiederum keine Wachstumsunterschiede zwischen diesen Linien fest. Als wir jedoch eine Kreuzung von +/+ x +/+ getrennt zu einer Kreuzung von -/- x -/- gehalten haben, stellten wir leichte Wachstumsunterschiede im Alter von 3 Monaten fest, die sich wahrscheinlich verstärken würden, wenn man den Fisch weiter wachsen ließe. Diese Unterschiede – einige größer, andere kleiner – waren unabhängig vom Lepr genotyp. - (Bagivalu Lakshminarasimha et al., 2021)
Fischwachstum – Werden Zebrafische fettleibig?
Eine Frage stellt sich, wenn man sieht, dass Leptin- oder Lepr-LOF-Tiere NICHT fettleibig sind: Werden Zebrafische überhaupt fettleibig? Und was für Effekte der Fettleibiugkeit können wir sehen?
Hier gibt es eine Veröffentlichung die zeight, dass überfütterte Zebrafische mehr Fett einlagern (Oka et al., 2010). Die Autoren zeigten auch eine erhöhte Lipidansammlung in der Leber, einen Anstieg der Plasmatriglyceride sowie konservierte Expressionsprofile zwischen viszeralem Fischfett und Säugetierfett. Allerdings ist Leptin, ein Hauptgen für die Expression und Sekretion im Fettgewebe, im Fettgewebe von Zebrafischen nicht vorhanden!
Deshalb haben wir den Fisch ab dem 5. Tag mit einer langfristigen adipösen Diät überfüttert und dies mit einer kalorienreduziertem Diät verglichen. Darüber hinaus erlaubten wir kalorienreduzierten Fischen, zu drei Wachstumszeitpunkten einen Ausgleich zu schaffen: zu Beginn ihres Lebens (1 Monat, in dem sie noch exponentiell wachsen), nach drei Monaten, in denen sie linear wachsen, und nach neun Monaten, in denen die Zebrafischwachstum ein Plateau erreicht. Wir stellten die Hypothese auf, dass eine Nahrungsmittelrestriktion früh in der Entwicklung der Fische eventuelle Endophänotypen der Fettleibigkeit verschlimmern würde.
Wir fanden heraus, dass Zebrafische zu allen drei Zeitpunkten im Wachstum aufholten und laut Jobling (2010) nicht durch kompensatorisches Wachstum. Das Gewicht-Längen-Verhältnis von Zebrafischen bei adipöser Ernährung zeigt ein positives allometrisches Wachstum auf, was bedeutet, dass Fische zunächst in der Länge wachsen und erst bei erreichen des Wachstumsplateaus anfangen in die Breite zu wachsen, d. h. fettleibig werden - ähnlich wie Menschen nur das Fische noch bis weit nach der Pubertät in der länge wachsen. In diesem älteren Stadium haben wir auch Hinweise auf Hepatosteatose und Hyperglykämie bei den fettleibigen Fischen gefunden, insgesamt waren die fettleibigen Fische jedoch weitgehend gesund. - (Leibold et al., 2022)
In gewisser Weise warf diese Studie mehr Fragen auf, als sie beantwortete. Wenn sich die Fettablagerung bei Fischen während des Alterns unterscheidet – ist es dann von entscheidender Bedeutung, wenn Lepr-LOF-Fische auf Adipositas getestet werden? Mäuse und Menschen zeigen Anzeichen einer schweren, früh einsetzenden krankhaften Fettleibigkeit (weit vor der Pubertät), Fische nicht. Allerdings zeigte die Gruppe von Herman Spaink, dass Leptin-b-LOF-Fische im Alter von 1–2 Jahren fettleibig werden (He et al., 2021). Wie verändert sich die Fettablagerung mit zunehmendem Alter der Fische? Natürlich ist das ein bekanntes Phänomen bei Menschen - die Fettverteilung ändert sich beim Altern.
Hinzu kommt - was sind eigneltich die Komorbiditäten oder Endophänotypen von Fettleibigkeit? Das Stoffwechselsyndrom umfasst viele Komorbiditäten, aber das Problem ist die Penetration – nicht alle Menschen leiden an diesen Komorbiditäten. Bisher haben wir die Linienunterschiede zwischen Zebrafischstämmen noch nicht untersucht. In einer kürzlich durchgeführten Studie untersuchte Helene Volkoff zwei verschiedene Zebrafischlinien und stellte fest, dass die Expression von wichtigen Energiehomöostase-Genen doch recht unterschiedlich war zwischen den Linien (London und Volkoff, 2019).
Die Rolle von Leptin in der Reproduktionsbiologie von Fischen
Bei der Charakterisierung der Agrp- und Lepr-LOF-Linien stellten wir fest, dass diese Fische einen ziemlich starken Rückgang der Hypophysen-Genexpression aufwiesen, insbesondere in der Fortpflanzungsachse (fsh, lh). Allerdings fanden wir keine offensichtlichen Reproduktions-Phänotypen. Dieses Ergebnis steht in krassem Gegensatz zu Daten aus dem Labor von Yonathan Zohar, die zeigen, dass Gnrh-LOF-Fische, die ebenfalls keinen offensichtlichen reproduktiven Phänotyp haben ganz normale Hypophysen-Genexpression aufweisen. Das heißt das diese Fische zentral, also im ZNS den Verlust des Gnrh Genes kompensieren (Marvel et al. 2018 & Marvel et al. 2019). In unserem Fall sehen fehlt Agrp bzw. Lepr die beide auf dem Gnrh Signalweg liegen, wir sehen auch keinen reproduktiven Phänotyp, jedoch einen starken Rückgang der Genexpression in der Hypophyse, dem Haupteffektor vonm Gnrh/Agrp/Lepr auf dieser Achse durch Exprimierung und Ausschüttung von FSH/LH. Bei den Lepr-LOF-Fischen haben wir bisher herausgefunden, dass sich die Fische zwar gut vermehren, wenn sie direkt aus den Haltungstanks geholt werden, dass sie jedoch eine Verzögerung der Reifung zeigen, wenn alle reifen Eizellen zuerst Paarungen gelegt werden und wir die Maturierung nach der Paarung verfolgen. Nach einer Paarung haben wir also in Lepr -/- Fischen gesehen, dass die Eizellen verzögert reifen und viel früher Anzeichen von Atresie zeigen. Atresie ist der Mechanismus der Resorption von Eizellen und ihre biologische Relevanz ist nicht ganz klar, aber es könnte sich um einen Mechanismus handeln, der die Eizellen bei Fischen relativ frisch hält, bis sich für den einzelnen Fisch eine Paarungsmöglichkeit ergibt, oder um ein Mittel zur Energieresorption. Dieser Vorgang findet auf natürliche Weise statt. Wenn weibliche Zebrafische von männlichen nach einer Paarung getrennt werden reifen die Oozyten circa 8 Tage lang bis der gonadosomatische Index ein Plateau erreicht, das heißt das die Gonaden nun nicht weiter wachsen. Zu diesem Zeitpunkt beginnt ein langsamer Zyklus kontinuierlicher Resorption und Reifung der sich in Balance hält. Lepr-LOF-Weibchen beginnen den Resorptionsprozess etwa 4 Tage früher und es ist bisher unklar warum. Darüber hinaus haben wir Mithilfe eines In-vitro-Tests zum Abbau von Keimbläschen (GVBD) gezeigt, dass exogenes Leptin und Lepr LOF einen Einfluss auf GVBD haben, also den letzten Schritt bei der Reifung der Oozyten. - (Bagivalu Lakshminarasimha et al., 2022).
Parallel zu unserer Studie zeigte Monika Schmitz in ihrem Labor, dass Lepr-LOF-Tiere für ein anderes Allel subfertil sind und deutlich schlechtere Leistungen erbringen als Wildtyp-Kontrollen. Sie zeigt weiterhin, dass dieser Effekt durch die Injektion von humanem Choriongonadotropin (hCG) außer Kraft gesetzt werden kann, was darauf hindeutet, dass der Effekt in ihren Händen primär zentral ist (Tsakoumis et al., 2022).